MỞ ĐẦU
Họ Bồ hòn (Sapindaceae Juss.) có khoảng 140 chi với khoảng 1500 loài, phân bố ở hầu hết các vùng nhiệt đới và cận nhiệt đới, một số có ở vùng ôn đới,... Việt Nam có 25 chi, 73 loài, 1 phân loài và 6 thứ, phân bố rải rác khắp cả nước. Giá trị tài nguyên của họ Bồ hòn khá quan trọng cả về khoa học lẫn giá trị sử dụng.
Các công trình nghiên cứu về hệ thống phân loại họ Bồ hòn được bắt đầu từ thế kỷ XVII, qua các công trình của C. Linnaeus (1753), A. J. Jussieu (1789), A. P. De Candolle (1824), C. L. Blume (1847), Miquel (1859),... Sau đó là hàng loạt tác giả với một số quan điểm khác nhau: G. Bentham & J. D. Hooker (1862), L. Radlkofer (1890, 1932), H. Scholz (1964), J. Muller & P. W. Leenhouts (1976), A. Takhtajan (1987, 1997).
Ở Việt Nam, cũng có những tác giả người nước ngoài nghiên cứu họ Bồ hòn từ rất sớm như J. Loureiro (1790), L. Pierre (1894-1895), H. Lecomte (1912), F. Gagnepain (1950). Bên cạnh đó là công trình của các tác giả người Việt Nam ngày càng nhiều và bổ sung thêm taxon và tư liệu cho hệ thực vật Việt Nam như Lê Khả Kế (1974), Phạm Hoàng Hộ (1970, 1992, 2000), Trần Đình Đại (1984), Trần Kim Liên (1993, 1994, 2003), Phan Kế Lộc và cs. (2001), Hà Minh Tâm và cs. (2003-2007),... Ngoài ra, có một số công trình về giá trị sử dụng của họ Bồ hòn như Võ Văn Chi (1991, 1997, 2003, 2004), Trần Công Khánh (1992), Nguyễn Ngọc Chính và cs. (1996), Đỗ Huy Bích và cs. (2004),...
Các nước lân cận với Việt Nam có các công trình nghiên cứu phân loại họ Bồ hòn, như D. Brandis (Ấn Độ) (1874), C. A. Backer & R. C. Bakhuizen f. (Inđônêxia) (1965), C. Y. Wu (Trung Quốc) (1977), H. S. Lo & T. Chen (Trung Quốc) (1985), F. Adema et al. (Malesiana) (1994), P. C. Welzen (Thái Lan) (1999),...
Thực vật chí Việt Nam, tập 12 – họ Bồ hòn (Sapindaceae Juss.) là công trình phân loại đầy đủ và có hệ thống, được biên soạn trên cơ sở hàng chục năm nghiên cứu. Việc điều tra thu thập mẫu tiêu bản và tư liệu được tiến hành khắp lãnh thổ Việt Nam, đặc biệt các vùng có điều kiện cho sự phân bố các loài họ Bồ hòn. Đồng thời, đã nghiên cứu tiêu bản và tư liệu ở các phòng tiêu bản trong nước như Viện Sinh thái và Tài nguyên sinh vật (HN), Trường Đại học Khoa học Tự nhiên – Đại học Quốc gia Hà Nội (HNU), Viện Sinh học Nhiệt đới – Tp. Hồ Chí Minh (VNM), Viện Dược liệu (HNPM), Viện Điều tra quy hoạch rừng (HNF), Trường Đại học Dược Hà Nội (HNIP) và Phòng Tiêu bản thực vật – Viện Thực vật Kunming – Trung Quốc (KUN) cũng như quan hệ hợp tác quốc tế để trao đổi thông tin.
Tập sách dày 358 trang, trình bày những nội dung cho một công trình hệ thống phân loại họ Bồ hòn ở Việt Nam. Giới thiệu đặc điểm hình thái các đại diện ở Việt Nam, kiểm kê các hệ thống phân loại và lựa chọn hệ thống cho nghiên cứu phân loại họ Bồ hòn ở Việt Nam. Phần chủ yếu là các khoá định loại taxon. Mỗi taxon được trình bày danh pháp, synonym, đặc điểm hình thái, mẫu chuẩn, sinh học và sinh thái, phân bố trong và ngoài nước, giá trị sử dụng, các nhận xét (nếu có). Bên cạnh đó là 82 hình vẽ, 40 trang ảnh màu; các bảng tra tên khoa học và Việt Nam, tài liệu tham khảo; tóm tắt tiếng Anh.
Trong công trình có tham khảo và trích dẫn tư liệu, hình vẽ, ảnh màu của các tác giả khác.
Cây tự dưỡng, sống lâu năm (trừ Cardiospermum đôi khi chỉ sống 1 năm), với dạng sống chủ yếu là cây thường xanh hoặc rụng lá một phần, ít khi rụng lá hoàn toàn trong một giai đoạn ngắn (Boniodendron, Schleichera). Kích thước cây rất thay đổi nhưng thường là cây gỗ cao tới 20-30 m hoặc cây bụi (chủ yếu ở Allophylus), ít khi là gỗ lớn cao tới 40-50 m (Amesiodendron, Pometia) và hiếm khi có dạng cây thảo leo (Cardiospermum); đường kính gốc hiếm khi vượt quá 1 m. Thân cây mọc thẳng đứng (trừ Cardiospermum và một số loài ở Allophylus), phân nhiều cành, một số có bạnh gốc (Pometia pinnata, Schleichera oleosa,...). Cành nhỏ có lỗ vỏ (bì khổng, bì khẩu). Phần non và các bộ phận của hoa có lông đơn, ít khi có lông hình sao mọc xen kẽ (Dimocarpus, Harpullia,...) hoặc lông tuyến (Dodonaea, Harpullia, Schleichera), hiếm khi có lông phân nhánh (Litchi) hoặc nhẵn hoàn toàn.
Lá luôn mọc cách theo kiểu xếp xoắn (trừ Litchi và Glenniea thỉnh thoảng có lá mọc đối ở gốc cụm hoa).
Dạng lá kép lông chim một lần (chẵn hoặc lẻ) hoặc kép chân vịt một lần (Allophylus và Delavaya), hiếm khi có lá kép 2 lần (Cardiospermum và Koelreuteria) hoặc có lá đơn (Dodonaea). Cuống lá tròn hoặc gần tròn, ít khi có mép hoặc có cánh (Guioa, Lepisanthes).
Lá chét mọc đối hoặc mọc cách, hình thái thay đổi từ hình trứng đến hình bầu dục hoặc trứng ngược, ít khi tròn hoặc có những hình dạng khác; chóp lá nhọn đến có đuôi, ít khi tù hoặc lõm; mép nguyên đến có răng cưa, hiếm khi có mép xẻ thùy (Cardiospermum); gốc cân đến lệch, nhọn đến tù, ít khi tròn hoặc hình tim; hai mặt không hoặc có lông với mức độ và các dạng lông khác nhau, ở một số chi mặt dưới có lỗ tuyến hoặc chùm lông ở nách gân bên (Allophylus, Dimocarpus, Glenniea và Koelreuteria); gân lá chét là gân hình lông chim; gân con hình lưới; cuống lá chét ngắn, hiếm khi không có.
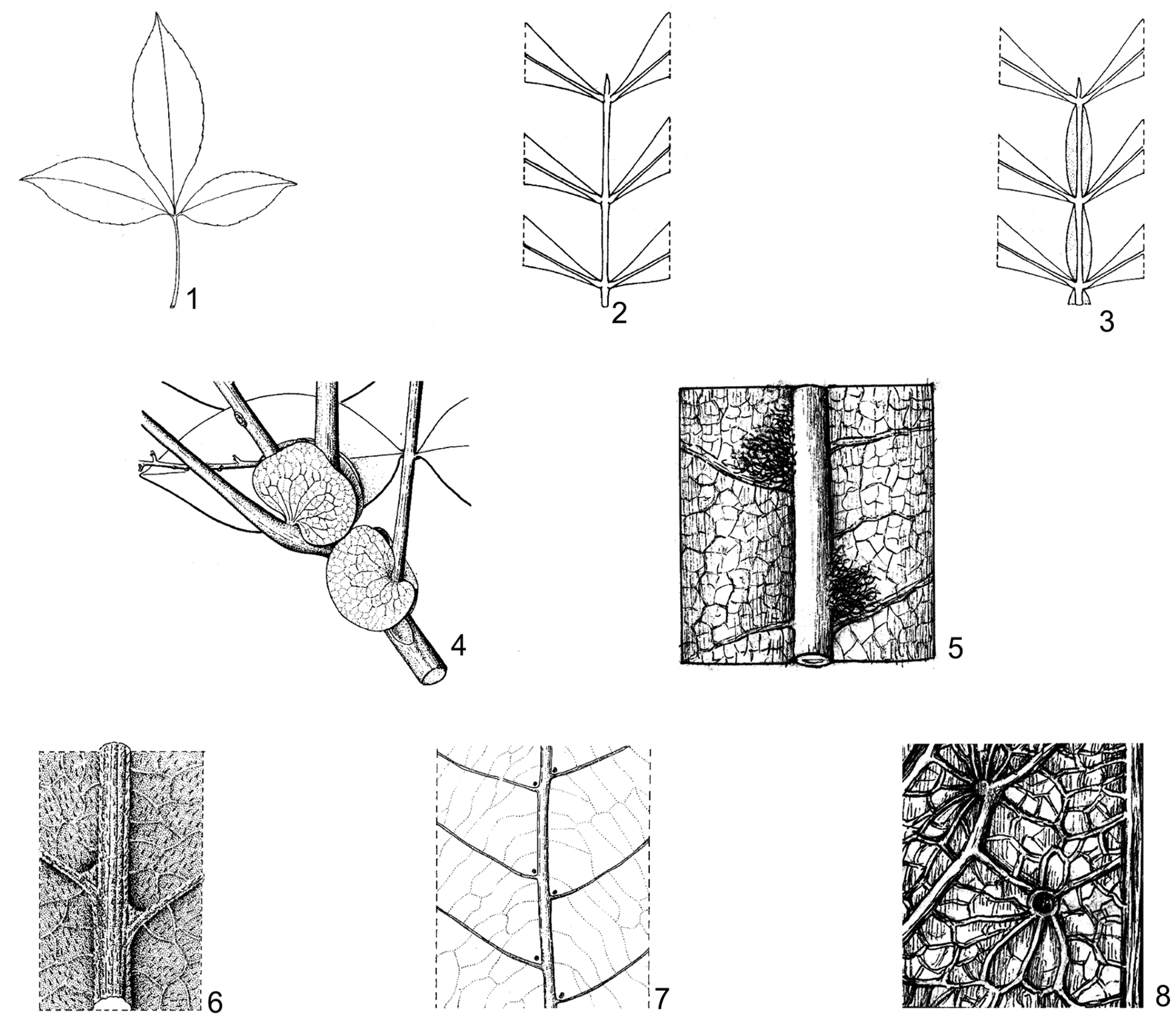
Hình 1. Một số đặc điểm về lá
1. lá kép chân vịt (Allophylus cobbe var. velutinus Corner); 2. lá kép lông chim chẵn, trục lá không có cánh (Guioa diplopetala (Hassk.) Radlk.); 3. lá kép lông chim chẵn, trục lá có cánh (Guioa pleuropteris (Blume) Radlk.); 4. lá kèm giả (Lepisanthes fruticosa (Roxb.) Leenh.); 5. chùm lông ở nách gân bên (Allophylus livescens Gagnep.); 6. lỗ tuyến ở gốc gân bên (Mischocarpus pentapetalus (Roxb.) Radlk.); 7. lỗ tuyến ở nách gân bên (Dimocarpus longan Lour.); 8. lỗ tuyến ở rải rác phiến lá (Xerospermum noronhianum (Blume) Blume) (1-4, 6, 7: theo Welzen, 1999; 5, 8: hình H. M. Tâm, 2006)
Lỗ tuyến có ở mặt dưới lá chét ở một số chi, gồm 2 loại: Một loại tạo thành lỗ sâu ở gốc gân bên (Arytera, Guioa, Mischocarpus,...); loại thứ 2 tạo thành lỗ nông ở nách gân bên hoặc ở rải rác phiến lá (Dimocarpus, Nephelium, Xerospermum, Pometia,...). Cho đến nay, các thông tin về lỗ tuyến ở họ Bồ hòn chỉ được biết rất ít qua các công trình của R. W. Ham (1977) và Welzen (1990, 1997) là các “domatia” không tiết dịch, xung quanh thường không có mô cứng nâng đỡ, còn chức năng của lỗ tuyến vẫn chưa được làm sáng tỏ. Ngoài ra, một số loài thuộc chi Lepisanthes có tuyến chìm (nốt màu trắng) ở phiến lá.
Lá kèm chỉ có ở chi Cardiospermum, nhưng sớm rụng.
Lá kèm giả có ở chi Pometia và Lepisanthes. Đó là đôi lá chét dưới cùng của lá kép không có cuống, đính ở gốc cuống lá kép, có hình thái và kích thước khác với những lá chét phía trên, trông giống như lá kèm.
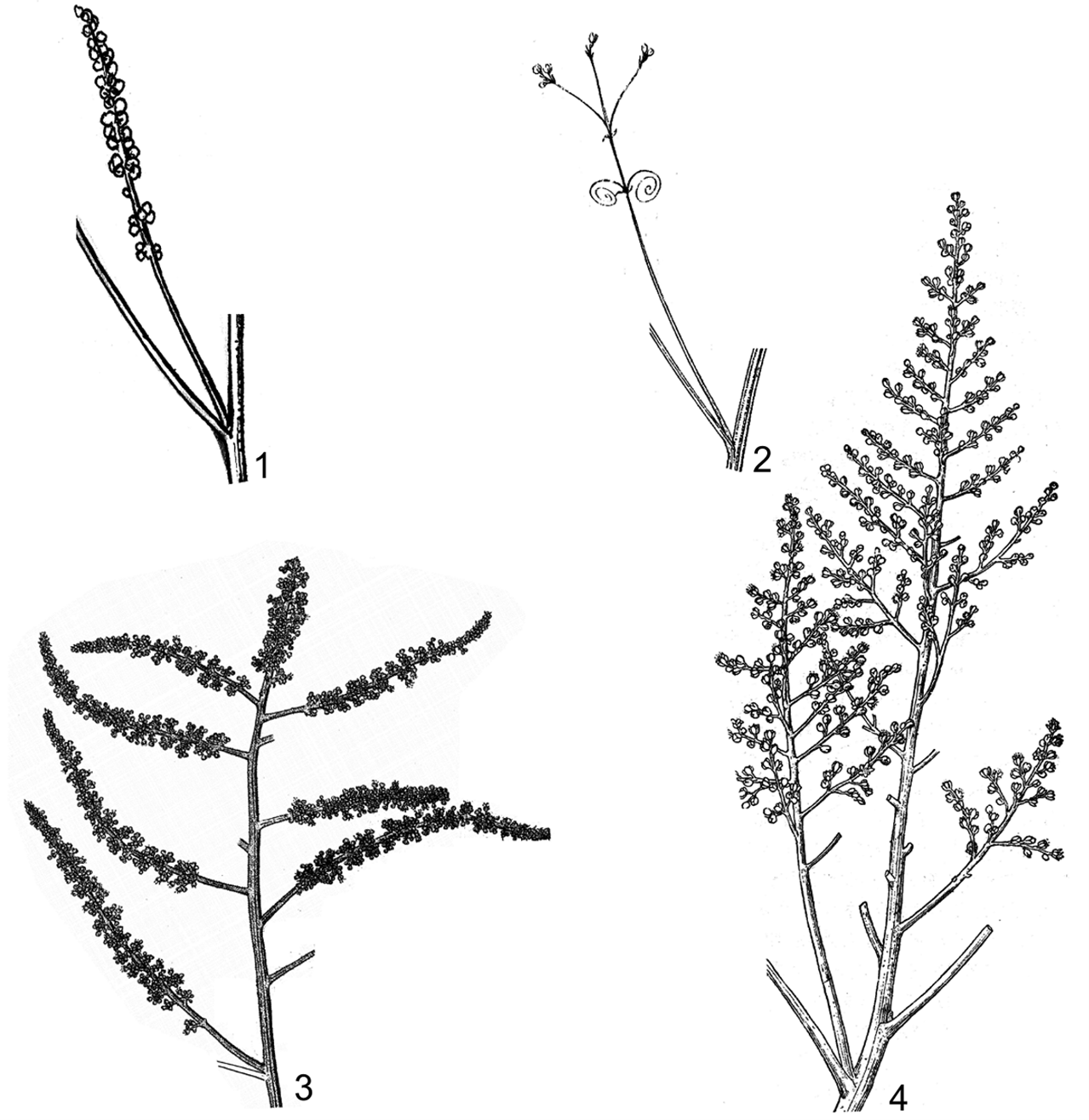
Hình 2. Một số hình thái cụm hoa
1. cụm hoa chùm ở nách lá, không phân nhánh (Allophylus hayatae Gagnep.); 2. cụm hoa chùy ở nách lá, có tua cuốn (Cardiospermum halicacabum L.); 3. cụm hoa chùy ở đỉnh, phân nhánh một lần (Glenniea philippinensis (Radlk.) Leenh.); 4. cụm hoa chùy ở đỉnh, phân nhánh hai lần (Sapindus saponaria L.) (1: hình H. M. Tâm, 2006; 2, 3: theo Welzen, 1999; 4: theo H. S. Lo & T. Chen, 1985)
Hầu hết hình chùy, đôi khi hình chùm (Allophylus, Blighia). Thường mọc ở nách lá gần đỉnh cành hoặc ở đỉnh cành, ít khi mọc ở phần thân hoặc cành không mang lá, đơn vị cơ sở là các xim 2 ngả, hiếm khi là xim 1 ngả (Guioa) hoặc ngù (Boniodendron), có lông. Lá bắc và lá bắc nhỏ hình tam giác, hình mũi mác hoặc hình đường,… ở chi Cardiospermum hai nhánh đầu tiên của cụm hoa biến thái thành tua cuốn.
Hầu hết có hoa đơn tính (cùng gốc hoặc khác gốc) (1), hiếm khi có hoa lưỡng tính (Dodonaea), hoa đều hoặc không đều.
Hoa lưỡng tính: Bao phấn mở, có hạt phấn; bộ nhụy phát triển thành quả.
Hoa đực: Bộ nhị phát triển (chỉ nhị dài, bao phấn mở và chứa hạt phấn), bộ nhụy không phát triển (bầu tiêu giảm và thui sau khi hoa nở, thường không có vòi nhụy), những hoa này chỉ có chức năng của hoa đực. [hình 3 (1-3)].
Hoa cái: Bộ nhụy phát triển và tạo quả (bầu phát triển, có vòi nhụy và núm nhụy), bộ nhị không phát triển (chỉ nhị ngắn hơn so với hoa đực cùng loài, bao phấn không mở và không chứa hạt phấn). Thoạt nhìn, hoa này rất dễ nhầm với hoa lưỡng tính. [hình 3 (4-6)].
Đài rời đến hợp, mẫu 4-5(-7), mặt ngoài có lông. Nếu đài rời thì lá đài không đều nhau, xếp lợp (Allophylus, Lepisanthes, Sapindus,...); nếu đài hợp thì các thùy đài thường đều nhau và xếp van (Koelreuteria, Litchi, Mischocarpus,…).
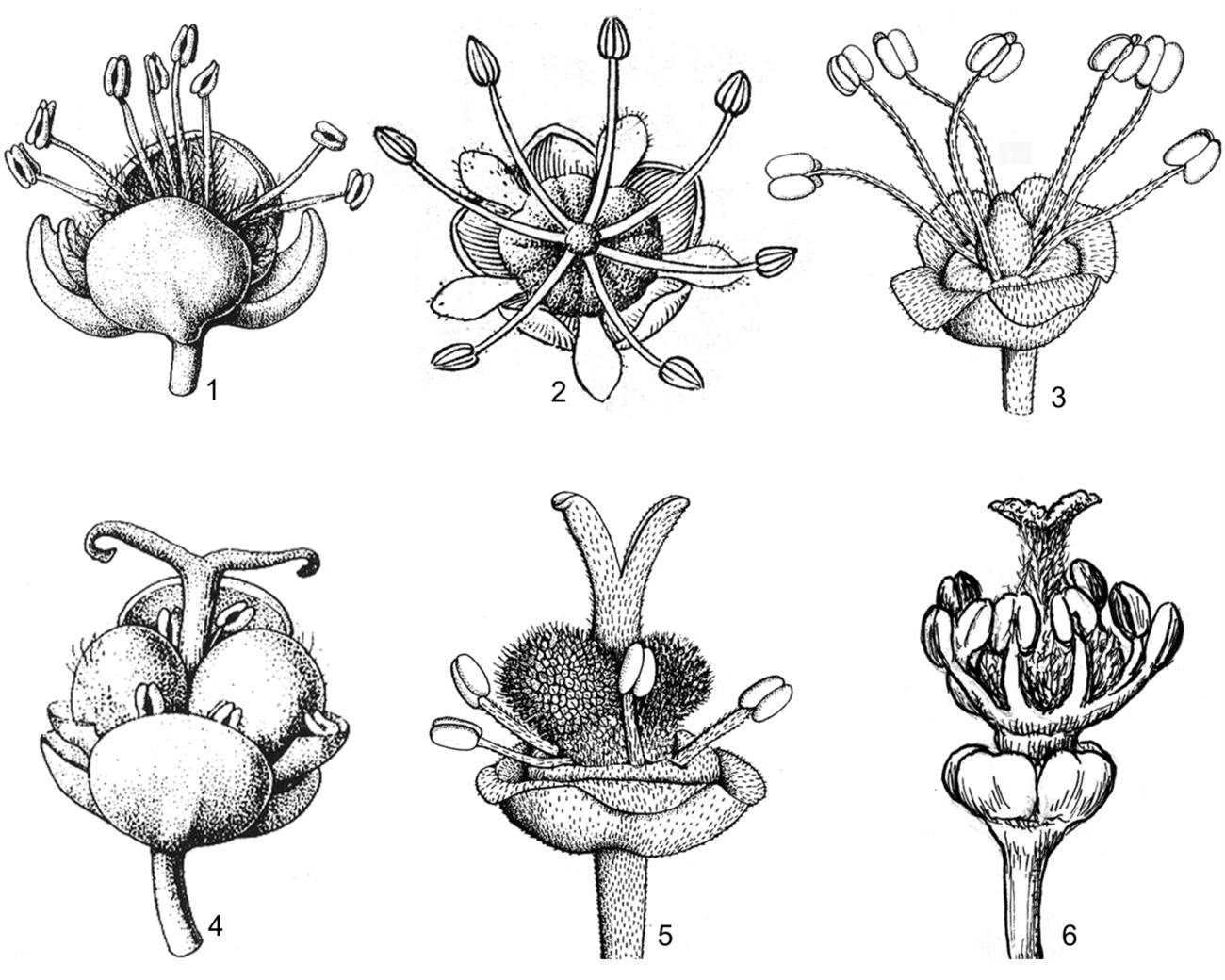
Hình 3. Một số đặc điểm về hoa
1. hoa đực với đài rời (Allophylus cobbe var. velutinus Corner); 2. hoa đực với đài hơi hợp, có 5 cánh hoa (Dimocarpus longan Lour.); 3. hoa đực có đài hợp, không có cánh hoa (Litchi chinensis Sonn.); 4. hoa cái không có cột nhụy, không có cột nhị-nhụy (Allophylus cobbe var. velutinus Corner); 5. hoa cái có cột nhụy (Litchi chinensis Sonn); 6. hoa cái có cột nhị-nhụy (Allophylus viridis Radlk.) (1, 3-5: theo Welzen, 1999; 2: theo H. S. Lo & T. Chen, 1985; 6: hình H. M. Tâm, 2006)
Tràng có sự giảm dần về số lượng cũng như kích thước cánh hoa, từ các đại diện có 4-5(-7) cánh hoa phát triển, mặt trong có vảy hoặc tai (Allophylus, Cardiospermum, Sisyrolepis, Boniodendron, Delavaya,...) đến các trường hợp có cánh hoa tiêu giảm và dao động từ 0-6, hầu hết không có vảy và tai (Dimocarpus, Mischocarpus, Nephelium) và cuối cùng là các đại diện hoàn toàn không có tràng (Dodonaea, Glenniea, Litchi, Schleichera). Cánh hoa có lông, hiếm khi nhẵn hoàn toàn. (hình 4).
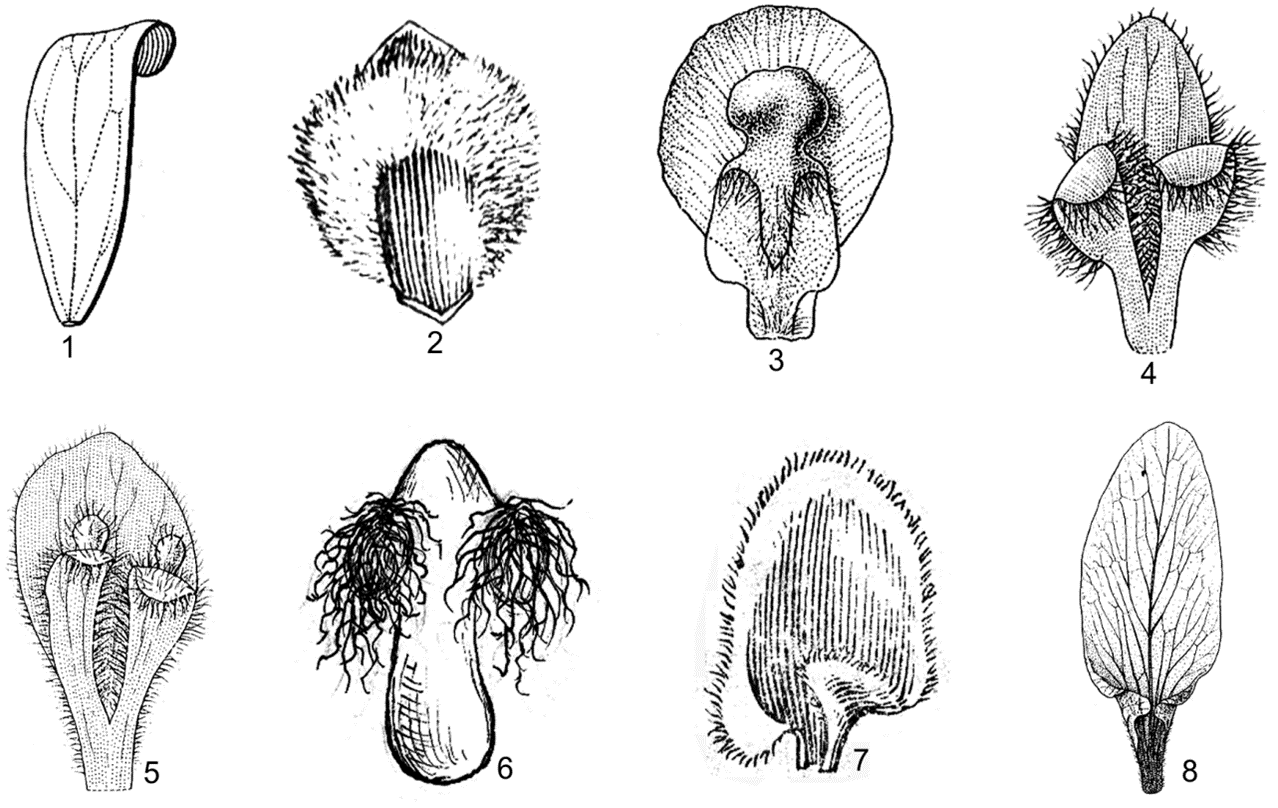
Hình 4. Một số đặc điểm hình thái cánh hoa
1. cánh hoa nhẵn hoàn toàn (Harpullia cupanioides Roxb.); 2. cánh hoa có một vảy nguyên, rậm lông (Pavieasia annamensis Pierre); 3. cánh hoa có một vảy nguyên, vảy có mào (Cardiospermum halicacabum L.); 4. cánh hoa có một vảy xẻ 2 thùy, không có mào (Guioa diplopetala (Hassk.) Radlk.); 5. cánh hoa có vảy xẻ 2 thùy, mỗi thùy có 1 mào (Guioa pleuropteris (Blume) Radlk.); 6. cánh hoa có vảy xẻ 2 thùy rất rậm lông (Allophylus livescens Gagnep.); 7, 8. cánh hoa có 2 tai ở gốc (7. Lepisanthes senegalensis (Poir.) Leenh.; 8. Harpullia arborea (Blanko) Radlk.) (1, 3-5, 8: theo Welzen, 1999; 2, 7: theo H. S. Lo & T. Chen, 1985; 6: hình H. M. Tâm, 2006)
Triền tuyến mật ở ngoài nhị, có ở hầu hết các chi (trừ hoa đực ở Dodonaea không có triền tuyến mật hoặc nếu có ở trong nhị); hình vành khuyên hoặc hình bán khuyên, một số hình chén nông, lượn sóng ở mặt trên hoặc xẻ sâu thành thùy rời nhau (Allophylus), ít khi có các hình thái khác, không hoặc có lông. [hình 5 (1-5)].
Cột nhị nhụy: Hình trụ, chỉ có ở một số loài thuộc chi Allophylus.
Cột nhụy: Hình trụ, có ở các chi Amesiodendron, Cardiospermum, Delavaya, Harpullia, Litchi, Mischocarpus,...
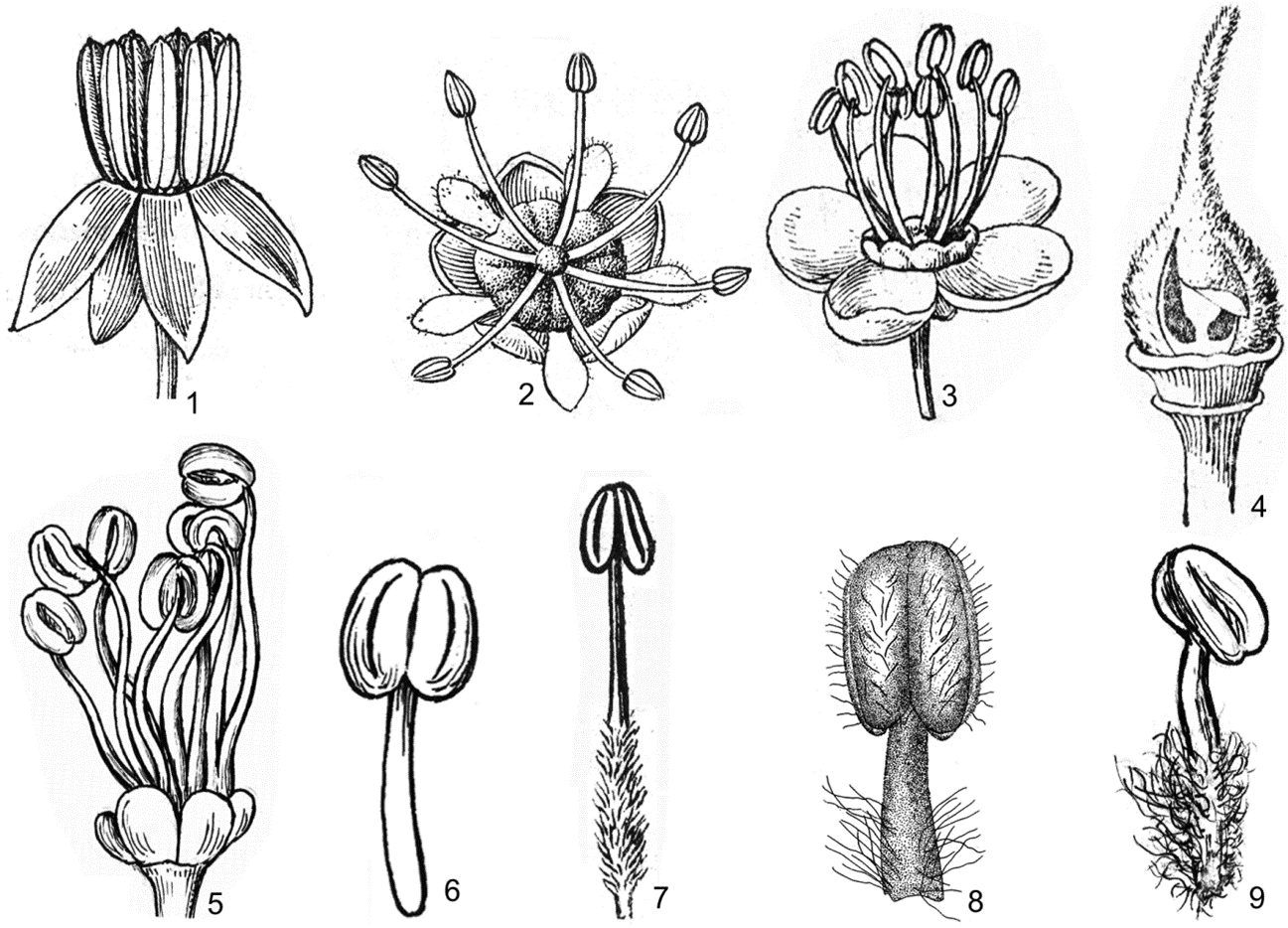
Hình 5. Một số đặc điểm về triền tuyến mật và nhị
1. không có triền tuyến mật, chỉ nhị rất ngắn so với bao phấn (Dodonaea viscosa (L.) Jacq.); 2, 3. triền tuyến mật hình vành khuyên, chỉ nhị dài hơn bao phấn (2. Dimocarpus longan Lour.; 3. Delavaya toxocarpa Franch.); 4. triền tuyến mật hình cốc (Pavieasia annamensis Pierre); 5. triền tuyến mật xẻ thành thùy rời nhau (Allophylus livescens Gagnep.); 6. nhị không có lông, bao phấn đính gốc (Mischocarpus sundaicus Blume); 7. chỉ nhị có lông, bao phấn đính gốc, không lông (Lepisanthes senegalensis (Poir.) Leenh.); 8. chỉ nhị và bao phấn có lông, bao phấn đính gốc (Koelreuteria paniculata Laxm.); 9. chỉ nhị có lông, bao phấn đính lưng, không lông (Allophylus hayatae Gagnep.) (1-4, 7: theo H. S. Lo & T. Chen, 1985; 5, 6, 9: hình H. M. Tâm, 2006; 8: theo F. G. Mey. 1976)
Bộ nhị có mặt ở cả hoa đực và hoa cái, nhưng ở hoa cái chỉ nhị ngắn hơn ở hoa đực, bao phấn không có hạt phấn và không mở; số nhị trong một hoa từ (5-)7-8(-13) nhị; đính ở bên trong triền tuyến mật (trừ Dodonaea); ở hoa đực nhị thường thò ra khỏi tràng. Chỉ nhị rời nhau, có lông hoặc không có lông. Bao phấn hình trứng hoặc hình thận; đính ở gốc hoặc thỉnh thoảng đính lưng ở sát gốc; mở bên theo chiều dọc bao phấn, ít khi mở hướng vào trong; trên bề mặt hiếm khi có lông. Hạt phấn có dạng đơn đẳng cực hoặc gần đẳng cực, hiếm khi tồn tại dạng tứ tử khi hạt phấn chín. Kích thước hạt phấn giữa 20-30 mm, hiếm khi chỉ có 13 mm (Cubilia) hoặc lên đến 74 mm (Cardiospermum). [hình 5 (6-9)].
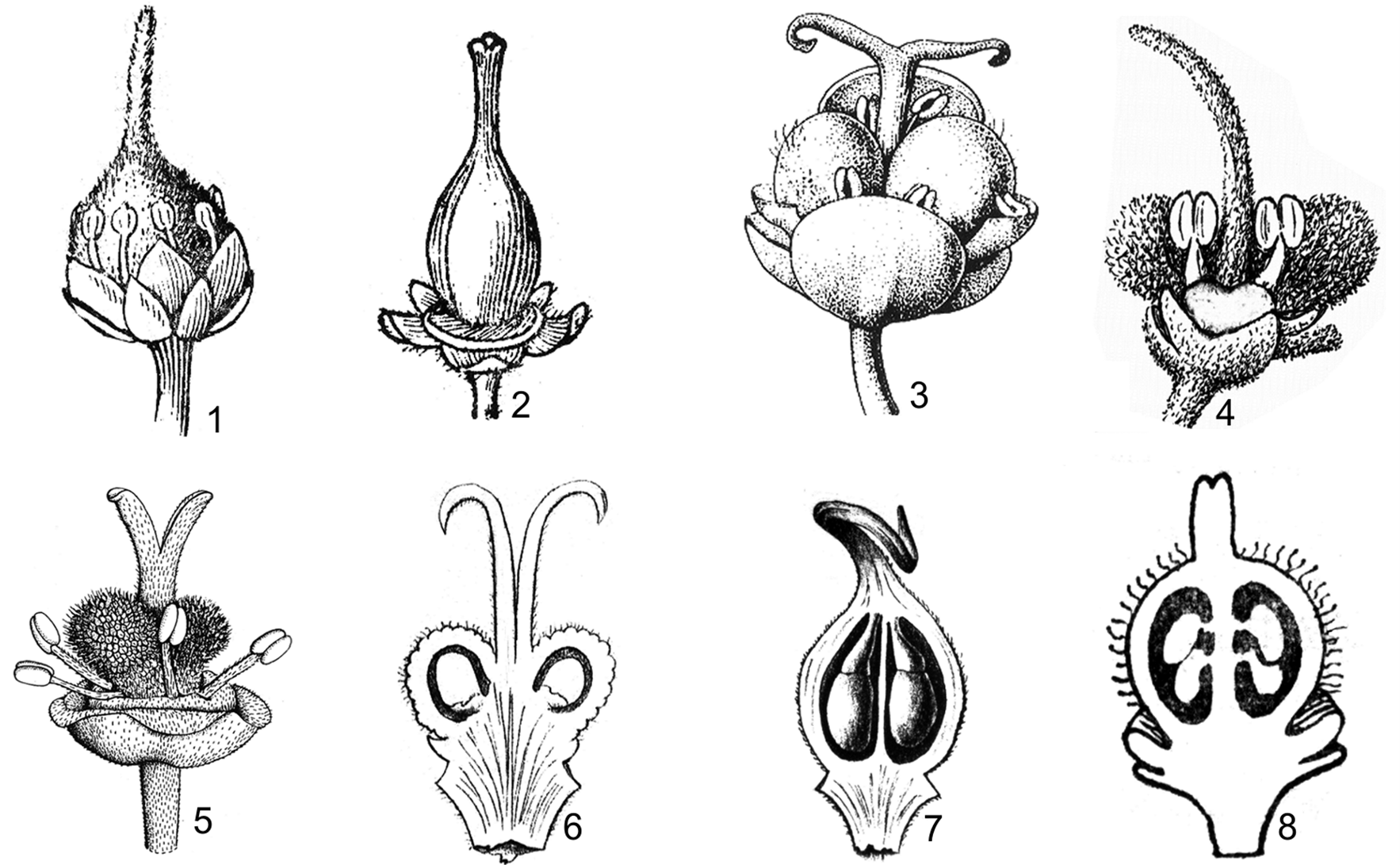
Hình 6. Một số đặc điểm về bộ nhụy
1. bầu không có thùy, rậm lông; vòi nhụy ở đỉnh bầu; núm nhụy nguyên (Pavieasia annamensis Pierre); 2. bầu 3 ô, hơi có thùy, nhẵn; vòi nhụy ở đỉnh bầu; núm nhụy hơi chia thùy (Sapindus saponaria L.); 3. bầu 2 ô, xẻ 2 thùy, có lông thưa; vòi nhụy ở đáy bầu; núm nhụy xẻ 2 thùy dài (Allophylus cobbe var. velutinus Corner); 4. bầu 2 ô, xẻ 2 thùy, rậm lông; vòi nhụy ở đáy bầu; núm nhụy nguyên (Pometia pinnata Forst & Forst. f.); 5. bầu 2 ô, xẻ 2 thùy, có nốt sần và lông; vòi nhụy ở đáy bầu; núm nhụy xẻ 2 thùy (Litchi chinensis Sonn); 6. mỗi ô bầu chứa 2 noãn rủ xuống (Boniodendron parviflorum (Lecomte) Gagnep.); 7. mỗi ô bầu chứa 1 noãn rủ xuống (Harpullia cupanioides Roxb.); 8. mỗi ô bầu chứa 1 noãn hướng lên (Nephelium cuspidatum var. bassacence (Pierre) Leenh.)
(1, 2, 6: theo H. S. Lo & T. Chen, 1985; 3, 5: theo Welzen, 1999; 4: hình H. M. Tâm, 2006; 7, 8: theo Pierre, 1894-1895)
Bộ nhụy có ở cả hoa đực và hoa cái, nhưng ở hoa đực bộ nhụy tiêu giảm và không tạo quả. Tất cả các đại diện đều có bầu thượng; gồm các lá noãn hợp hoàn toàn (Boniodendron, Dodonaea, Paranephelium,…) hoặc chỉ hợp một phần ở gốc bầu và vòi nhụy (Allophylus, Dimocarpus, Litchi, Nephelium,…); có (1-)2-3(-4) ô; mặt ngoài bầu có lông (Allophylus, Arytera, Boniodendron, Lepisanthes, Pavieasia,…) hoặc có nốt sần và lông dày đặc (Dimocarpus, Litchi, Nephelium, Paranephelium, Xerospermum,...), hiếm khi nhẵn hoàn toàn; mỗi ô của bầu chứa 2-3 noãn, hiếm khi chỉ chứa 1 noãn (ở phân họ Dodonaeoideae) hoặc chỉ chứa 1 noãn (ở phân họ Sapindoideae). Noãn không có cuống, đính trực tiếp trên giá noãn (trừ Koelreuteria). Vòi nhụy có hình trụ, đính ở đỉnh bầu (khi nguyên hoặc chỉ hơi có thùy) hoặc đính ở phần đế hoa giữa các thùy bầu (khi bầu xẻ thùy sâu); ngắn hơn hoặc dài hơn chiều cao của bầu, hiếm khi không có vòi nhụy (ở một số loài thuộc chi Lepisanthes). Núm nhụy nguyên (Delavaya, Koelreuteria, Lepisanthes, Pavieasia,...), có khía (Nephelium, Guioa) hoặc xẻ thùy sâu (Allophylus, Cardiospermum, Dimocarpus, Litchi, Mischocarpus, Nephelium, Paranephelium, Sapindus,...). (hình 6).
Gồm 2 loại chính là quả mở (quả nang) và quả không mở (quả hạch và một số dạng quả khác), nhưng hình thái rất phức tạp: quả nguyên (Sisyrolepis, Schleichera,...), có thùy (Guioa, Lepisanthes,...) đến xẻ thùy sâu thành từng phần riêng biệt([1]) (Dimocarpus, Litchi, Nephelium,…), ít khi quả có cánh (Dodonaea, Boniodendron, Zollingeria).
Hầu hết quả đạt tới kích thước tối đa ngay từ lúc hạt vẫn chưa phát triển hoàn toàn, nhưng sau một thời gian khá dài thì quả mới chín. Ở những loài có áo hạt, thì áo hạt phát triển đồng thời với hạt. Trong giai đoạn hạt phát triển, quả có màu xanh, vỏ cứng và có hàm lượng tanin cao. Khi chín hầu hết có màu đỏ, màu vàng hoặc màu tím, với độ đậm nhạt thay đổi; vỏ quả rất thay đổi: vỏ mỏng và phồng to (Cardiospermum, Koelreuteria), mềm như da (Dimocarpus, Litchi, Xerospermum,...) hoặc hóa gỗ (Amesiodendron, Arytera, Guioa, Paranephelium,...); mặt ngoài có lông (Harpullia, Lepisanthes,…), có nốt sần (Litchi, Nephelium, Xerospermum,…) hoặc gai mềm (Nephelium, Paranephelium, Sisyrolepis,…), ít khi nhẵn hoàn toàn (Pometia, Sapindus,…).
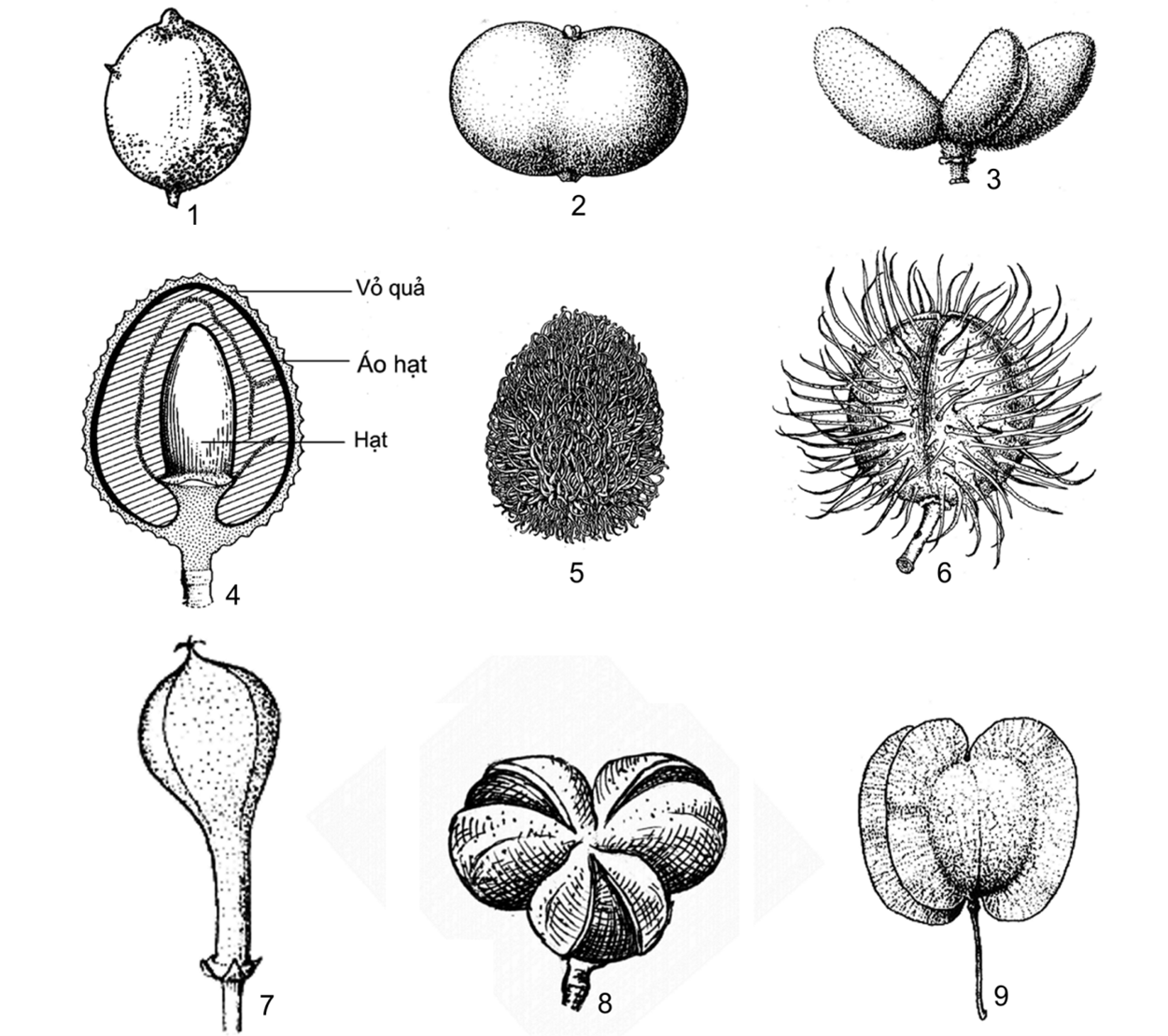
Hình 7. Một số đặc điểm về quả
1. quả hạch, không chia thùy, vỏ có vài gai cứng (Schleichera oleosa (Lour.) Oken); 2. quả hạch, hơi có thùy (Lepisanthes fruticosa (Roxb.) Leenh.); 3. quả hạch, xẻ sâu thành 3 thùy (Lepisanthes rubiginosa (Roxb.) Leenh.); 4. thùy quả bổ dọc (Litchi chinensis Sonn); 5. quả hạch, vỏ có gai mềm (Nephelium lappaceum L.); 6. quả nang, không chia thùy, vỏ có gai mềm (Sisyrolepis muricata (Pierre) Leenh.); 7. quả nang, hơi có thùy, nửa trên gần hình cầu, nửa dưới hình trụ (Mischocarpus sundaicus Blume); 8. quả nang, xẻ 3 thùy (Amesiodendron tienlinense H. S. Lo); 9. quả có cánh (Dodonaea angustifolia L. f.) (1, 8: hình H. M. Tâm, 2006; 2-7, 9: theo Welzen, 1999)
Hạt không có nội nhũ, không có cuống (trừ Koelreuteria), thường có hình cầu, hình trứng hoặc hình bầu dục, hiếm khi có hình tháp (Zollingeria); màu sắc biến đổi từ xám sẫm đến đen; thường không có lông.
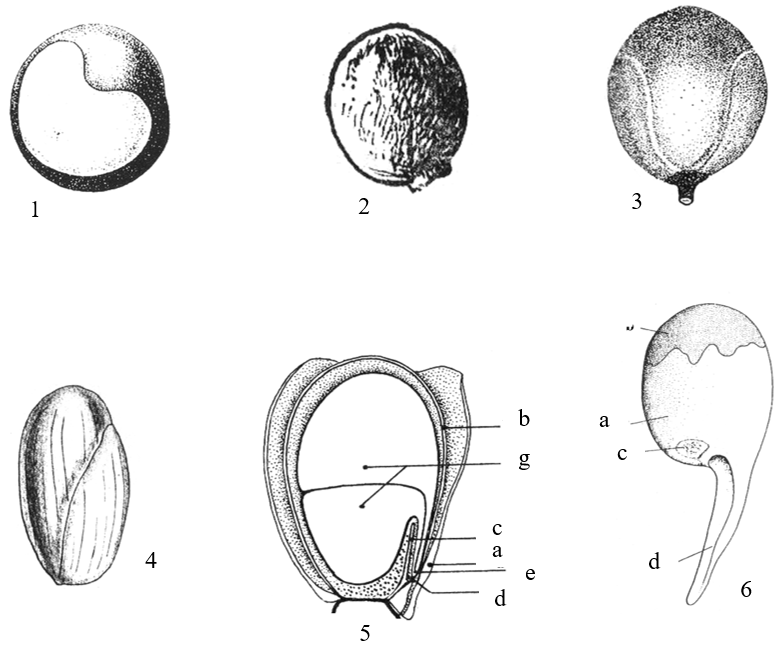
Hình 8. Một số đặc điểm về hình thái hạt
1. hạt nhẵn bóng, rốn hạt hình tim (Cardiospermum halicacabum L.); 2. hạt nhẵn bóng, rốn hạt tròn (Delavaya toxocarpa Franch.); 3. hạt có cuống (Koelreuteria paniculata Laxm.); 4. hạt đã tách vỏ, rốn hạt rất nhỏ (Nephelium lappaceum L.); 5. hạt bổ dọc (Arytera littoralis Blume: a-áo hạt, b-vỏ hạt, c-nếp gấp trong của vỏ hạt, d-lỗ noãn, e-rễ phôi, g-lá mầm); 6. hạt có cuống noãn giả (Mischocarpus: a-áo hạt, b-vỏ hạt, c-rốn hạt, d-cuống noãn giả) (1: theo Welzen, 1999; 2: theo H. S. Lo & T. Chen, 1985; 3: theo F. G. Mey. 1976; 4: theo Pierre, 1894; 5, 6: theo R. W. Ham, 1977)
Áo hạt (áo hạt giả, cơm quả, thịt quả, tử y) có ở một số chi, bao bên ngoài hạt, gồm hai loại: áo hạt được hình thành từ rốn hạt và không dính vào vỏ hạt (Arytera, Blighia, Dimocarpus, Guioa, Harpullia, Litchi, Mischocarpus, Pometia, Schleichera), áo hạt được hình thành từ vỏ hạt và dính chặt vào vỏ hạt (Harpullia, Nephelium, Xerospermum); một số chi có cuống noãn giả (phần phụ của áo hạt kéo dài gắn hạt vào vách quả: Guioa, Mischocarpus).
Vỏ hạt dai hoặc mềm, ít khi hóa gỗ (Sapindus); rốn thường hình trứng đến tròn, ít khi hình tim (Cardiospermum) hoặc hình dải (Sapindus).
Phôi dày, cong hình cung, đôi khi có nếp gấp hoặc xoắn ốc. Lá mầm dày và thường xếp chồng lên nhau (khi đó phôi uốn cong) hoặc ở bên cạnh nhau (khi đó phôi có gờ) nhưng nhiều khi xen kẽ giữa hai dạng trên.
Hạt được phát tán chủ yếu nhờ con người và động vật (khỉ, dơi, cáo, lười, gấu, vẹt,...), đôi khi nhờ gió và nước (Dodonaea, Pometia, Sapindus).
Typus: Sapindus L.
Họ Bồ hòn (Sapindaceae Juss.) trên thế giới có khoảng 140 chi với khoảng 1500 loài, phân bố ở hầu hết các vùng nhiệt đới và cận nhiệt đới, một số có ở vùng ôn đới; chủ yếu ở những nơi ẩm ướt và có nhiều ánh sáng trong rừng thứ sinh, như mép rừng, bờ suối, ven lối đi; trong mọi điều kiện thổ nhưỡng; thường ở độ cao dưới 1000(-3600) m. Ở Việt Nam, hiện gặp 25 chi, 74 loài, 1 phân loài và 6 thứ (trong đó có 5 loài hiện biết không đầy đủ). Cho đến nay, ngoài một số cây trồng: Nhãn (Dimocarpus longan) được trồng gần như khắp cả nước, Vải (Litchi chinensis) được trồng gần khắp miền Bắc và một số tỉnh miền Trung và Chôm chôm (Nephelium lappaceum) được trồng gần khắp miền Nam), đã thống kê được 49/63 tỉnh và thành phố có mặt các cây hoang dại thuộc họ Bồ hòn. Trong đó, tỉnh có nhiều loài nhất là Đồng Nai (với 23 loài hoang dại và 4 loài là cây nhập trồng), Lâm Đồng (18 loài hoang dại và 2 loài nhập trồng), Lạng Sơn (16 loài hoang dại và 2 loài nhập trồng), Ninh Bình (15 loài hoang dại và 2 loài nhập trồng),… Các tỉnh có ít loài nhất là: Điện Biên (1 loài hoang dại: Hoạt bi nam (Harpullia cupanioides) và 2 loài nhập trồng), Đồng Tháp (1 loài hoang dại: Chôm chôm (Nephelium lappaceum) và 2 loài nhập trồng), Cần Thơ (1 thứ hoang dại: Chôm chôm rừng (Nephelium lappaceum var. pallens) và 2 loài nhập trồng), Hậu Giang (1 thứ hoang dại: Chôm chôm hậu giang (Nephelium cuspidatum var. bassacense) và 2 loài nhập trồng),… 16/63 tỉnh chưa phát hiện thấy loài hoang dại là Bắc Ninh, Hải Dương, Hưng Yên, Thái Bình, Nam Định, Phú Yên, Bình Phước, Long An, Tiền Giang, Bến Tre, An Giang, Trà Vinh, Vĩnh Long, Sóc Trăng, Bạc Liêu, Cà Mau.
Mùa hoa từ tháng 1-6 hàng năm, quả chín sau khi hoa nở khoảng 4-6 tháng (trừ một số loài mùa quả chín kéo dài tới mùa hoa năm sau: Schleichera oleosa). Hầu hết các loài thụ phấn chéo nhờ côn trùng, hiếm khi nhờ gió (Dodonaea). Ở những cây cùng gốc phổ biến là sự chín không đồng thời của nhị và nhụy (biệt giao) với các pha khác nhau (đầu tiên là pha của hoa đực nở trước, sau khi hoa đực rụng đi thì xuất hiện pha của hoa cái. Tuy nhiên đôi khi sự tách biệt giữa 2 pha là không rõ ràng).
Sự nảy mầm: Hạt có tuổi thọ ngắn và không có trạng thái ngủ. Trong khoảng 1 tuần đầu tiên, nếu điều kiện thích hợp thì tỷ lệ nảy mầm ở hạt rất cao, càng về sau thì tỷ lệ nảy mầm càng thấp, nhất là ở những loài hạt chứa nhiều dầu béo. Khi nảy mầm, rễ mầm, cuống lá mầm và trụ dưới lá mầm phồng lên thành dạng ống phá vỡ vỏ rốn hạt (nơi có rễ phôi), sau đó vỏ hạt sẽ mở theo một đường dài ở vỏ bên. Rễ con thường mọc xung quanh lớp vỏ. Sau đó cuống lá mầm, trụ dưới lá mầm và rễ kéo dài ra. Dần dần, lá mầm chuyển thành màu xanh.
Cây con lúc mới nảy mầm có đặc điểm hình thái khác nhiều so với cây trưởng thành. Hầu hết có lá kép 3, mọc đối, cuống hơi có cánh, mép lá chét có khía tròn đến có răng cưa (hiếm khi nguyên), không có lỗ tuyến và gai thịt ở mặt dưới lá chét, kích thước lá chét khác nhiều so với cây trưởng thành.
Họ Bồ hòn (Sapindaceae Juss.) ở Việt Nam có số loài không lớn, nhưng có nhiều giá trị quan trọng. Cho đến nay, đã xác định được 10 loài thuộc họ Bồ hòn mới chỉ gặp ở Việt Nam là Ngoại mộc cánh ngắn (Allophylus brachypetalus), Ngoại mộc chùm ngắn (A. brachystachys), Ngoại mộc nam (A. cochinchinensis), Ngoại mộc nâu (A. fuscus), Ngoại mộc hayata (A. hayatae), Ngoại mộc hoa thưa (A. laxiflorus), Ngoại mộc tái (A. livescens), Lân hùng bà nà (Lepisanthes banaensis), Chét (Sapindus chrysotrichus) và Nhãn rừng (Sapindus sonlaensis), 1 loài được ghi trong Sách Đỏ Việt Nam (2007) là Bông mộc (Sinoradlkofera minor). Các tài liệu về giá trị sử dụng đã ghi nhận 3 loài được trồng rộng rãi làm cây ăn quả (loài Nhãn - Dimocarpus longan và Vải - Litchi chinensis được đánh giá là những cây ăn quả chủ đạo ở miền Bắc, loài Chôm chôm - Nephelium lappaceum được trồng phổ biến ở miền Nam), 37 loài cây cho gỗ, 28 loài được dùng làm thuốc, 14 loài cho dầu béo, 11 loài chứa saponin, 7 loài chứa flavonoid, một số loài được trồng làm cây bóng mát,... với tổng số 49 loài (thuộc 25 chi) đã và đang được khai thác về giá trị kinh tế. Ngoài ra, một số loài còn được trồng làm cây tiên phong trong việc phục hồi rừng trên núi đá vôi như Dầu choòng (Delavaya toxocarpa); cây chủ nuôi cánh kiến đỏ như Dấu dầu (Schleichera oleosa); cây bóng mát có tính chất trang trí như A kê (Blighia sapida), Hoạt bi nam (Harpullia cupanioides), Tầm phong gỗ (Koelreuteria bipinnata), Lát núi (Koelreuteria paniculata), Bồ hòn (Sapindus saponaria),...
Trước khi họ Bồ hòn (Sapindaceae Juss.) được công bố, C. Linnaeus (1753) đã công bố một số chi và loài sau này được xếp vào họ Bồ hòn, trong đó chi Cupania được xếp vào nhóm 5 nhị với 1 vòi nhụy (pentandria monogynia); chi Allophylus xếp vào nhóm 8 nhị với 1 vòi nhụy (octandria monogynia); Paullinia, Cardiospermum, Sapindus được xếp vào nhóm 8 nhị với 3 vòi nhụy (octandria trigynia),… Sau này đã cùng một số tác giả khác còn công bố một số chi mới nhưng vẫn sắp xếp giống như trên.
A. J. Jussieu (1789) đã xếp 21 chi vào họ này và đặt tên là Sapindi và chia thành 3 nhóm:
I. Cánh hoa kép (ngày nay được xác định là mặt trong cánh hoa có vảy), một số ít có cựa (8 chi): Cardiospermum, Paullinia, Sapindus,...
II. Cánh hoa có lông (9 chi): Schmidelia, Euphoria, Melicocca, Cossignia,...
III. Các chi có họ hàng gần gũi với các nhóm trên (4 chi): Matayba, Enourea, Cupania và Pekea.
Vào năm 1811, tác giả đã chỉnh lý lại tên gọi thành Sapindaceae. Về sau, các công trình nghiên cứu về họ này đều lấy tên là Sapindaceae Juss. (1789).
Sau khi hệ thống phân loại của A. J. Jussieu được công bố, đã có một số hệ thống phân loại họ Bồ hòn ra đời, như các hệ thống của A. P. De Candolle (1824), C. L. Blume (1847), Miquel (1859),... Tuy các hệ thống này được xây dựng vào thời điểm khoa học về phân loại nói chung và phân loại thực vật nói riêng vẫn chưa thực sự phát triển, các dẫn liệu để xây dựng hệ thống vẫn chưa thật sự đầy đủ, về mặt danh pháp còn nhiều thiếu sót,... Nhưng đây là những căn cứ quan trọng về ý tưởng xây dựng hệ thống và đặc điểm nhận biết một số taxon. Cho đến nay, có nhiều hệ thống phân loại họ Bồ hòn ra đời, nhưng có thể tóm tắt thành 2 quan điểm chính như sau:
Quan điểm thứ nhất: Đặt các họ Aceraceae Juss. 1789, Melianthaceae Link, 1831 và Staphyleaceae Lindl. 1829 nằm trong họ Sapindaceae và chia họ này thành 5 tông. Đây là quan điểm của các tác giả G. Bentham & J. D. Hooker (1862) và W. P. Hiern (1875), cụ thể như sau: Trong hệ thống của G. Bentham & J. D. Hooker (1862), họ Sapindaceae gồm 73 chi, được chia thành 5 subordo (thực tế hiểu là tông) là: Sapindeae, Acerineae, Dodonaeae, Meliantheae và Staphyleae. Hệ thống của W. P. Hiern (1875) chỉ có một sự thay đổi đáng kể so với hệ thống của G. Bentham & J. D. Hooker là việc sửa lại thuật ngữ subordo thành tribus.
Quan điểm thứ hai: Đặt các họ Aceraceae Juss. 1789, Melianthaceae Link, 1831 và Staphyleaceae Lindl. 1829 là các họ độc lập. Họ Sapindaceae được chia thành 2 phân họ và các tông. Theo quan điểm này có các tác giả L. Radlkofer (1890, 1932), H. Scholz (1964), J. Muller & P. W. Leenhouts (1976), A. Takhtajan (1987, 1997), cụ thể như sau: Trong các hệ thống của L. Radlkofer (1890, 1932), họ Sapindaceae được chia thành 2 series (thực tế hiểu là phân họ) với 14 tribus. Trong hệ thống của H. Scholz (1964), tác giả đã chỉnh lý lại tên gọi và thay đổi vị trí 2 phân họ so với các hệ thống của L. Radlkofer. Hệ thống của J. Muller & P. W. Leenhouts (1976) là hệ thống được xây dựng trên cơ sở các hệ thống của L. Radlkofer, nhưng căn cứ vào các dẫn liệu mới, các tác giả đã nhập tông Aphanieae vào Lepisantheae, thay đổi trật tự của hầu hết các tông trong cả 2 phân họ và đề xuất một cách không chính thức 3 nhóm (A, B và C) trong phân họ Sapindoideae. Các hệ thống của A. Takhtajan (1987, 1997) về cơ bản giống với hệ thống của J. Muller & P. W. Leenhouts (1976), tác giả chỉ thay đổi vị trí phân loại của một số tông và loại bỏ 3 nhóm không chính thức (A, B và C) trong hệ thống của J. Muller & P. W. Leenhouts.
Các quan điểm về hệ thống phân loại họ Sapindaceae nêu trên được tóm tắt trong bảng 1 dưới đây:
Bảng 1. TÓM TẮT MỘT SỐ HỆ THỐNG CHÍNH ĐƯỢC SỬ DỤNG ĐỂ PHÂN LOẠI HỌ BỒ HÒN (SAPINDACEAE Juss.) TRÊN THẾ GIỚI
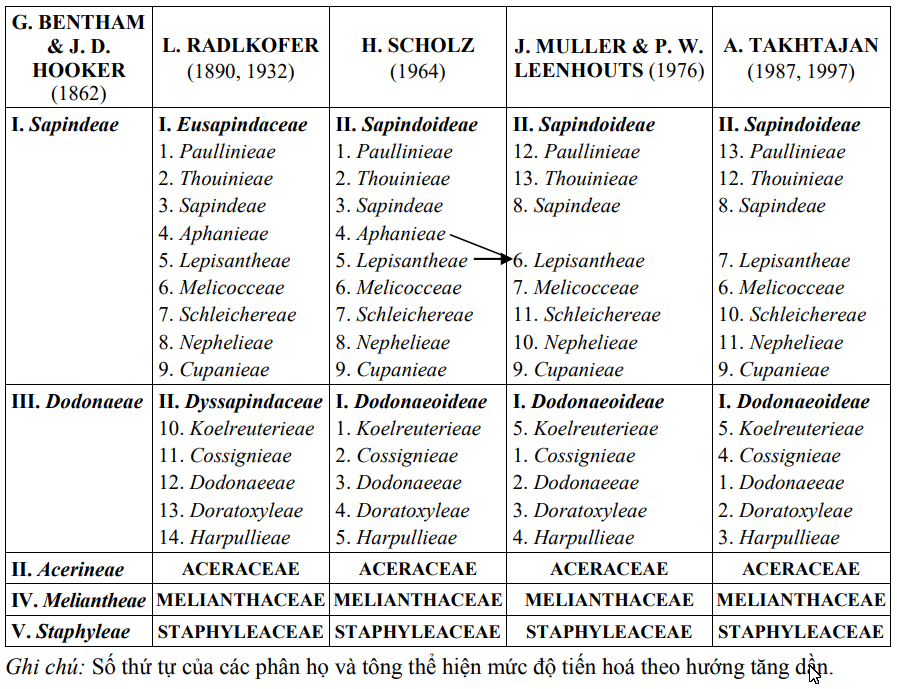
Ngoài các hệ thống nêu trên, còn một số công trình đề cập đến hệ thống phân loại họ này nhưng dưới dạng danh lục, đó là các công trình của J. C. Willis (1973) và D. J. Mabberley (1997). Các tác giả đã mô tả tóm tắt họ, giới thiệu đặc điểm nhận biết 2 phân họ và cung cấp một số thông tin về phân loại các chi,...
Các vùng lân cận Việt Nam có một số công trình nghiên cứu họ Bồ hòn theo các hệ thống khác nhau, như công trình nghiên cứu họ Bồ hòn ở vùng Tây Bắc và trung tâm Ấn Độ của D. Brandis (1874). Tác giả đã dựa vào hệ thống của G. Bentham & J. D. Hooker để xây dựng khoá định loại và mô tả 6 chi và 13 loài. C. A. Backer & R. C. Bakhuizen f. (1965) dựa vào hệ thống của L. Radlkofer (1932) để xây dựng khóa định loại và mô tả 26 chi với 37 loài thuộc họ Bồ hòn có ở đảo Java (Inđônêxia). Các tác giả Trung Quốc (1972) dựa vào hệ thống G. Bentham & J. D. Hooker để sắp xếp và giới thiệu đặc điểm nhận biết 24 loài thuộc họ Bồ hòn ở Trung Quốc. C. Y. Wu (1977) xây dựng khoá định loại và mô tả 18 chi, 24 loài, 2 phân loài và 9 thứ thuộc họ Bồ hòn có ở Vân Nam (Trung Quốc). N. P. Balakrisnan (1981) dựa vào hệ thống J. Muller & P. W. Leenhouts để xây dựng khoá định loại và mô tả 5 chi, 7 loài thuộc họ Bồ hòn có ở Jowai (miền Bắc Ấn Độ). K. Haridasan & R. R. Rao (1985) xây dựng khoá định loại và mô tả 8 chi và 10 loài thuộc họ Bồ hòn ở Meghalaya (Ấn Độ). H. S. Lo & T. Chen (1985), khi nghiên cứu hệ thực vật Trung Quốc đã xây dựng khóa định loại và mô tả 25 chi với 55 loài, 2 phân loài và 3 thứ thuộc họ Bồ hòn. Yap (1989) xây dựng khoá định loại và mô tả 17 chi, 49 loài và 11 thứ thuộc họ Bồ hòn ở Malaixia. Islam (1990) dựa vào hệ thống của L. Radlkofer (1932) để xây dựng khoá định loại và mô tả 4 chi và 4 loài thuộc họ Bồ hòn có ở Majuli (Ấn Độ). C. Y. Wu (1991) xây dựng khoá định loại và mô tả 17 chi, 23 loài và 1 phân loài thuộc họ Bồ hòn có ở Vân Nam (Trung Quốc). C. H. Chen (1993) xây dựng khoá định loại và mô tả 7 chi với 7 loài thuộc họ Bồ hòn ở Đài Loan. D. J. D. Puy & J. R. H. Telford (1993) xây dựng khoá định loại cho 6 chi, 3 loài và 1 phân loài thuộc họ Bồ hòn có ở Ôxtrâylia. F. Adema & al. (1994), khi nghiên cứu họ Bồ hòn ở vùng Malesiana đã mô tả 44 chi, 241 loài, 15 phân loài và 14 thứ. P. C. Welzen (1999) mô tả 23 chi, 40 loài, 3 phân loài và 3 thứ thuộc họ Bồ hòn ở Thái Lan. Phần lớn số tác giả này dựa theo hệ thống của J. Muller & P. W. Leenhouts (1976) và L. Radlkofer (1932).
Ở Việt Nam, công trình của J. Loureiro (1790) là công trình đầu tiên nghiên cứu hệ thực vật Việt Nam. Trong công trình này, tác giả đã mô tả 8 chi và 13 loài thuộc họ Bồ hòn, trong đó có 2 chi mới và 10 loài mới (hiện chỉ có 1 chi và 1 loài là tên chính thức, 3 loài biết vẫn chưa đầy đủ, còn lại là tên đồng nghĩa). Tác giả sắp xếp theo hệ thống của C. Linnaeus (1753).
L. Pierre (1894-1895) nghiên cứu hệ thực vật rừng Nam Bộ đã mô tả 20 chi với 44 loài và 5 thứ thuộc họ Bồ hòn, trong đó có 4 chi mới, 36 loài mới và 5 thứ mới (hiện chỉ có 3 loài được công nhận, 2 loài và 1 thứ là tên đồng nghĩa gốc, 7 loài không hợp luật, còn lại là các tên đồng nghĩa). Tác giả đã công nhận họ Sapindaceae là một taxon bậc họ riêng biệt, nhưng không đề cập đến hệ thống phân loại.
H. Lecomte (1912) nghiên cứu họ Bồ hòn ở Đông Dương đã mô tả 26 chi và 64 loài. Trong đó có 24 chi với 53 loài ở Việt Nam (hiện nay 7 chi và 36 loài đã trở thành các tên đồng nghĩa hoặc không hợp luật, 1 chi và 1 loài không thuộc họ Bồ hòn). Tác giả sắp xếp theo hệ thống của G. Bentham & J. D. Hooker (1862).
F. Gagnepain (1950) trong phần bổ sung cho Thực vật chí đại cương Đông Dương của Lecomte năm 1912, đã xây dựng khoá định loại cho 28 chi, xây dựng khoá định loại đến loài cho 28 chi, thống kê 103 loài (mô tả 56 loài chưa được mô tả trong công trình của H. Lecomte năm 1912) thuộc họ Bồ hòn ở Đông Dương. Trong số 26 chi và 89 loài ở Việt Nam có 6 chi và 44 loài trong công trình này hiện đã trở thành tên đồng nghĩa hoặc không hợp luật. Về mặt hệ thống, tác giả dựa vào hệ thống của G. Bentham &
J. D. Hooker (1862).
Phạm Hoàng Hộ (1970) trong "Cây cỏ miền Nam Việt Nam" đã xây dựng khoá định loại cho 17 chi, tóm tắt đặc điểm nhận biết 47 loài và 1 thứ thuộc họ Bồ hòn ở miền Nam. Trong đó 4 chi, 20 loài là tên đồng nghĩa, 5 loài và 1 thứ là tên không hợp luật. Tác giả không đề cập đến hệ thống phân loại.
Lê Khả Kế và cộng sự (1974) trong “Cây cỏ thường thấy ở Việt Nam” cũng xây dựng khóa định loại cho 18 chi và mô tả 28 loài thuộc họ Bồ hòn. Thực tế đây là công trình được trích dịch từ Thực vật chí đại cương Đông Dương nên cũng lặp lại những thiếu sót giống như công trình này.
Phạm Hoàng Hộ (1992) trong "Cây cỏ Việt Nam" đã bổ sung một số chi và loài, đưa tổng số chi và loài của họ Bồ hòn ở Việt Nam lên 26 chi với 66 loài, 2 phân loài và 3 thứ. Trong đó 1 chi và 1 loài (Melicoccus bijugatus) không có ở Việt Nam, 4 chi và 7 loài này là các tên đồng nghĩa. Về hệ thống phân loại, trong khoá tác giả coi Otophora, Erioglossum, Aphania, Lepisanthes là 4 chi độc lập (giống hệ thống Radlkofer, 1932) nhưng trong phần mô tả loài tác giả nhập tất cả 3 chi trên vào Lepisanthes (giống hệ thống Muller & Leenhouts, 1976). Năm 2000, trong "Cây cỏ Việt Nam" được tái bản, tác giả đã xây dựng khoá định loại cho 27 chi, tóm tắt đặc điểm nhận biết của 67 loài, 2 phân loài và 4 thứ. Trong đó 1 chi và 4 loài không có ở Việt Nam, 1 chi và 3 loài là tên đồng nghĩa, 1 loài là tên không hợp luật. Các chi Aphania, Erioglossum và Otophora được nhập vào Lepisanthes (giống hệ thống Muller & Leenhouts, 1976) nhưng giống như công trình của ông năm 1992, các taxon không sắp xếp theo trật tự của hệ thống này [tông Lepisantheae được thay thế bằng tông Aphanieae, chi Pavieasia được xếp vào phân họ Dodonaeoideae (trong tông Harpullieae), chi Crossonephelis (= Glenniea) xếp vào tông Nephelieae,...]. Cho đến nay, chúng tôi chưa thấy công trình nào có cách sắp xếp giống như các công trình của Phạm Hoàng Hộ.
Ngoài ra, ở Việt Nam còn có một số công trình nghiên cứu về họ Bồ hòn dưới hình thức là giáo trình (Phạm Hoàng Hộ, 1975; Võ Văn Chi và Dương Đức Tiến, 1978); cẩm nang tra cứu (Nguyễn Tiến Bân, 1997; Nguyễn Nghĩa Thìn, 1997) hoặc danh lục, bài báo phát hiện taxon mới ở Việt Nam (Trần Đình Đại, 1984; Trần Kim Liên, 1993, 1994, 2003; Phan Kế Lộc và cs., 2001; Hà Minh Tâm và cs., 2003-2007);...
Bên cạnh các công trình chuyên nghiên cứu về phân loại nêu trên, còn một số công trình đề cập đến giá trị tài nguyên của họ Bồ hòn ở Việt Nam như các công trình của Cục Điều tra quy hoạch rừng (nay là Viện Điều tra quy hoạch rừng) vào các năm 1971, 1978, 1981, 1982, 1986, 1988; Vũ Văn Chuyên (1976); Đỗ Tất Lợi (1977, 1995, 2003); Võ Văn Chi (1991, 1997, 2003, 2004); Trần Công Khánh & Phạm Hải (1992); Trần Đình Lý (1993); Đỗ Huy Bích và cs. (2004);...
Như vậy, có thể thấy rằng, các công trình nghiên cứu phân loại họ Bồ hòn ở Việt Nam, trong đó các công trình được coi là có hệ thống nhất của H. Lecomte (1912) và F. Gagnepain (1950) đến nay đã không còn phù hợp, các công trình của Phạm Hoàng Hộ chỉ mang tính chất giới thiệu về đặc điểm nhận biết các loài cây họ Bồ hòn ở Việt Nam. (Bảng 2). Vì vậy, việc chọn lọc và xây dựng một hệ thống cho họ Bồ hòn ở Việt Nam phù hợp với hiện tại là rất cần thiết.
Qua tìm hiểu các công trình nghiên cứu họ Bồ hòn (Sapindaceae Juss.), chúng tôi thấy mỗi tác giả đều có những quan điểm riêng của mình về việc lựa chọn hệ thống phân loại, nhưng số tác giả dựa vào quan điểm hệ thống G. Bentham & J. D. Hooker (1862) không nhiều và chủ yếu là những công trình nghiên cứu về họ này trong phạm vi hẹp. Bên cạnh đó, các công trình nghiên cứu dựa vào quan điểm hệ thống L. Radlkofer (1890, 1932) ngày càng được ủng hộ. Nếu như hệ thống G. Bentham & J. D. Hooker (1862) được sử dụng nhiều trong giai đoạn đầu khi mới ra đời, thì càng về sau các hệ thống của L. Radlkofer (1890, 1932) lại được sử dụng nhiều hơn. Trong các công trình gần đây, hầu hết các tác giả (nhất là những tác giả nghiên cứu những vùng lớn) đã dựa vào hệ thống L. Radlkofer (1890, 1932) trên cơ sở chấp nhận những bổ sung của J. Muller & P. W. Leenhouts (1976) để nghiên cứu họ Bồ hòn, nhưng quan điểm chia thêm các nhóm không chính thức (A, B, C) của J. Muller & P. W. Leenhouts lại tỏ ra không phù hợp với thực tế phân loại, điều đó chứng tỏ việc loại bỏ các nhóm này trong các hệ thống của A. Takhtajan là hoàn toàn hợp lý.
Bảng 2. CÁCH SẮP XẾP CÁC CHI THUỘC HỌ BỒ HÒN (SAPINDACEAE Juss.) Ở VIỆT NAM THEO F. GAGNEPAIN (1950) VÀ PHẠM HOÀNG HỘ (2000)

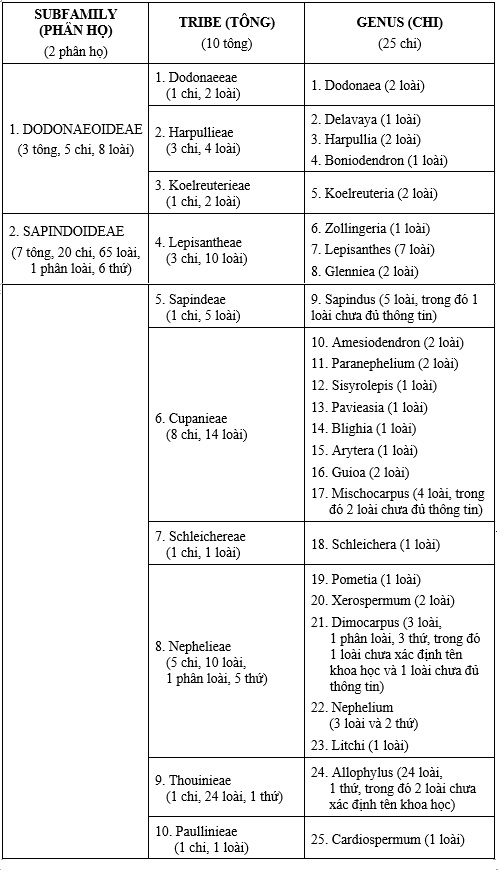
Sau khi phân tích và so sánh các hệ thống phân loại họ Bồ hòn (Sapindaceae), chúng tôi đã lựa chọn hệ thống của A. Takhtajan (1997) để sắp xếp các taxon thuộc họ Bồ hòn ở Việt Nam, vì hệ thống này được xây dựng trên cơ sở kế thừa có chọn lọc kết quả của các hệ thống L. Radlkofer (1890, 1932), J. Muller & P. W. Leenhouts (1976) – những hệ thống nghiên cứu về họ Sapindaceae trên phạm vi toàn thế giới và được đánh giá là tương đối hoàn chỉnh. Ngoài ra, tác giả đã cập nhật những thông tin về danh pháp, những dẫn liệu mới về phân loại, các dẫn liệu về hoá sinh học,... của các tác giả nghiên cứu về họ Bồ hòn trên thế giới để sắp xếp và giải thích mối quan hệ giữa các taxon. Đồng thời khắc phục được những mặt còn hạn chế trong các công trình kể trên (như việc loại bỏ các nhóm không chính thức A, B và C trong công trình của J. Muller & P. W. Leenhouts, sắp xếp lại vị trí của một số tông,...). Cho đến nay, cách sắp xếp của hệ thống này phù hợp với việc phân loại họ Sapindaceae của hầu hết các tác giả nghiên cứu về họ này trên thế giới và phù hợp với quan điểm phân loại họ Bồ hòn ở Việt Nam hiện nay, cho nên chúng tôi lựa chọn hệ thống này. Trên cơ sở hệ thống A. Takhtajan (1997), họ Bồ hòn ở Việt Nam được chia thành 2 phân họ, 10 tông và 25 chi, với 74 loài, 1 phân loài và 6 thứ (Bảng 3).
Bảng 3. TÓM TẮT HỆ THỐNG PHÂN LOẠI HỌ BỒ HÒN (SAPINDACEAE Juss.) Ở VIỆT NAM
(2 phân họ, 10 tông và 25 chi, với 73 loài, 1 phân loài và 6 thứ)
(Thường gặp trong các mục “Typus” và “Mẫu nghiên cứu”)
|
A
|
= Arnold Arboretum, Cambridge, USA.
|
|
B
|
= Botanisches Museum, Koeningin-Luise-Strasse, Berlin, Germany.
|
|
BM
|
= British Museum (Natural History), London, UK.
|
|
BO
|
= Herbarium Bogoriense, Bogor, Indonesia.
|
|
C
|
= Botanical Museum and Herbarium, Copenhagen, Denmark.
|
|
G
|
= Conservatoire et Jardin Botaniques, Geneve, Switzland.
|
|
VNM (HM)
|
= Herbarium, Institute of Tropical Biology, Ho Chi Minh City, Vietnam. (Phòng Tiêu bản thực vật, Viện Sinh học nhiệt đới, tp. Hồ Chí Minh).
|
|
HN
|
= Herbarium, Institute of Ecology and Biological Resources, Hanoi Vietnam. (Phòng Tiêu bản thực vật, Viện Sinh thái và Tài nguyên sinh vật).
|
|
HNF
|
= Herbarium, Forest Inventory and Planning Institute, Hanoi, Vietnam. (Phòng Tiêu bản thực vật, Viện Điều tra Quy hoạch rừng).
|
|
HNIP
|
= Herbarium, Hanoi University of Pharmacy, Vietnam.
(Phòng Tiêu bản thực vật, trường Đại học Dược, Hà Nội).
|
|
HNMM
|
= Herbarium, National Institute of Medicinal Materials, Hanoi, Vietnam. (Phòng Tiêu bản thực vật, Viện Dược liệu, Hà Nội)
|
|
HNU
|
= Herbarium, Hanoi National University, Hanoi, Vietnam (Phòng Tiêu bản thực vật, trường Đại học Khoa học Tự nhiên, Đại học Quốc gia Hà Nội).
|
|
K
|
= The Herbarium and Library, Royal Botanical Gardens, Kew, Surrey, UK.
|
|
KUN
|
= Kunming Botanical Institute, Academia Sinica, Kunming, Yunnan, China.
|
|
L
|
= Rijksherbarium, Nonnensteeg, Leiden, The Netherlands.
|
|
LE
|
= Botanical Institute Komarov, Leningrad (St. Petersbourg), RFR. (USSR).
|
|
LINN
|
= The Linnean Society of London, London, UK.
|
|
M
|
= Botanische Staatssamlung, Muenchen, Germany.
|
|
MO
|
= Missouri Botanical Garden, Missouri, USA.
|
|
NSW
|
= National Herbarium of New South Wales, Botanic Gardens, Sydney Australia.
|
|
NY
|
= The New York Botanical Garden, New York, USA.
|
|
P
|
= Museum National d' Histoire Naturelle, Paris, France.
|
|
PE
|
= Institute of Botany, Academia Sinica, Peiking, China.
|
|
TAI
|
= The Herbarium, Department of Botany, National Taiwan University, Taiwan.
|
|
U
|
= Botanical Museum and Herbarium, 106 Lange, Neiuwstraat, Utrecht, The Netherlands.
|
|
UC
|
= Herbarium of the University of California, Berleley, California, USA.
|
|
W
|
= Naturhistorique Museum, Wien, Austria.
|
| 1A. |
Mỗi ô của bầu chứa 2-3 noãn, hiếm khi chỉ có 1 noãn (khi đó noãn rủ xuống). Quả nang. (Subfam. 1. DODONAEOIDEAE) |
|
| 2A. |
Lá đơn. Cánh hoa 0. (Trib. 1. Dodonaeeae) |
1. DODONAEA |
| 2B. |
Lá kép. Cánh hoa 4-5 |
|
| 3A. |
Lá kép hình chân vịt hoặc kép hình lông chim chẵn một lần. Đài xếp lợp. Nhị không có lông. Hạt không có cuống. (Trib. 2. Harpullieae) |
|
| 4A. |
Lá kép hình chân vịt |
2. DELAVAYA |
| 4B. |
Lá kép hình lông chim chẵn |
|
| 5A. |
Mép lá chét nguyên. Quả không có cánh |
3. HARPULLIA |
| 5B. |
Mép lá chét có răng cưa. Quả có cánh |
4. BONIODENDRON |
| 3B. |
Lá kép hình lông chim lẻ hoặc kép hình lông chim chẵn hai lần. Đài xếp van. Nhị có lông. Hạt có cuống. (Trib. 3. Koelreuterieae) |
5. KOELREUTERIA |
| 1B. |
Mỗi ô của bầu chỉ chứa 1 noãn (noãn thẳng đứng hoặc hướng lên). Quả nang hoặc quả hạch (Subfam. 2. SAPINDOIDEAE) |
|
| 6A. |
Cây gỗ hoặc cây bụi. Cuống cụm hoa không có tua cuốn |
|
| 7A. |
Lá kép hình lông chim. Cụm hoa hầu hết phân nhánh. Hầu hết là cây gỗ |
|
| 8A. |
Hầu hết có cánh hoa; cánh hoa phát triển, luôn có vảy hoặc tai (trừ Zollingeria – khi đó quả có cánh). Bầu nguyên, hiếm khi xẻ thùy (Lepisanthes) |
|
| 9A. |
Quả không mở |
|
| 10A. |
Quả nguyên, có thùy hoặc có cánh, nếu xẻ thùy sâu thì dính nhau ở gốc. Hạt hiếm khi có lông, rốn hạt gần hình tròn (Trib. 4. Lepisantheae) |
|
| 11A. |
Quả có cánh. Cánh hoa không có vảy và tai |
6. ZOLLINGERIA |
| 11B. |
Quả không có cánh. Nếu có cánh hoa thì có vảy hoặc tai ở gốc |
|
| 12A. |
Cánh hoa 4-5(-7). Chỉ nhị có lông |
7. LEPISANTHES |
| 12B. |
Cánh hoa 0. Chỉ nhị không có lông |
8. GLENNIEA |
| 10B. |
Quả luôn xẻ thùy sâu, khi chín thùy tách rời nhau. Hạt có nhiều lông ở gốc, rốn hạt hình dải (Trib. 5. Sapindeae) |
9. SAPINDUS |
| 9B. |
Quả mở (Trib. 6. Cupanieae) |
|
| 13A. |
Lá chét không có lỗ tuyến |
|
| 14A. |
Quả có thùy rõ. Mép lá chét luôn có răng cưa |
10. AMESIODENDRON |
| 14B. |
Quả nguyên hoặc hơi có thùy. Mép lá chét nguyên hoặc có răng cưa |
|
| 15A. |
Vỏ quả có gai hoặc có nốt sần |
|
| 16A. |
Lá kép hình lông chim lẻ. Chỉ nhị không có lông |
11. PARANEPHELIUM |
| 16B. |
Lá kép hình lông chim chẵn. Chỉ nhị có lông |
12. SISYROLEPIS |
| 15B. |
Vỏ quả không có gai, không có nốt sần |
|
| 17A. |
Lá chét 5-13 đôi. Không có áo hạt |
13. PAVIEASIA |
| 17B. |
Lá chét 4-7 đôi. Có áo hạt |
14. BLIGHIA |
| 13B. |
Lá chét có lỗ tuyến ở gốc gân bên. |
|
| 18A. |
Cánh hoa 4-5, mặt trong có vảy. Quả có thùy rõ. |
|
| 19A. |
Cuống quả không có đài tồn tại. Hạt không có cuống noãn giả |
15. ARYTERA |
| 19B. |
Cuống quả có đài tồn tại. Hạt có cuống noãn giả |
16. GUIOA |
| 18B. |
Cánh hoa 0-5, gốc có 2 tai. Quả hơi có thùy |
17. MISCHOCARPUS |
| 8B. |
Không có cánh hoa hoặc nếu có thì cánh hoa tiêu giảm, không có vảy và tai. Bầu xẻ thùy tương ứng với số ô, hiếm khi nguyên (Schleichera). |
|
| 20A. |
Bầu và quả nguyên; vòi nhụy ở đỉnh bầu (Trib. 7. Schleichereae) |
18. SCHLEICHERA |
| 20B. |
Bầu và quả xẻ thùy; vòi nhụy ở giữa các thùy bầu (Trib. 8. Nephelieae) |
|
| 21A. |
Có lá kèm giả. Bầu có lông. Núm nhụy không chia thùy. Vỏ quả nhẵn |
19. POMETIA |
| 21B. |
Không có lá kèm giả. Bầu có nốt sần và lông. Núm nhụy chia 2-3 thùy. Vỏ quả luôn có nốt sần hoặc có gai |
|
| 22A. |
Đài xếp lợp, tồn tại ở quả. Hoa không có cột nhụy. Hạt hình cầu |
|
| 23A. |
Cánh hoa 4-5. Vỏ hạt thô ráp. Lá chét 1-2(-3) đôi, luôn mọc đối |
20. XEROSPERMUM |
| 23B. |
Cánh hoa 0-5(-6). Vỏ hạt nhẵn bóng. Lá chét 3-7 đôi, hiếm khi mọc đối |
21. DIMOCARPUS |
| 22B. |
Đài xếp van, không tồn tại ở quả. Hoa có cột nhụy. Hạt hình bầu dục |
|
| 24A. |
Lá chét thường có lỗ tuyến ở mặt dưới. Cánh hoa 0-5. Vỏ hạt thô ráp |
22. NEPHELIUM |
| 24B. |
Lá chét không có lỗ tuyến. Cánh hoa 0. Vỏ hạt nhẵn bóng |
23. LITCHI |
| 7B. |
Lá kép hình chân vịt (có 1, 3 hoặc 5 lá chét). Cụm hoa hiếm khi phân nhánh. Hầu hết là cây bụi (Trib. 9. Thouinieae) |
24. ALLOPHYLUS |
| 6B. |
Cây thảo. Cuống cụm hoa có tua cuốn (Trib. 10. Paullinieae) |
25. CARDIOSPERMUM |
(1) Cho đến nay, việc sử dụng thuật ngữ để chỉ một phần quả bị xẻ ra ở những loài thuộc họ Bồ hòn có quả xẻ thùy sâu vẫn chưa thống nhất. Ví dụ để chỉ một phần quả bị xẻ ra của 1 quả ở Nephelium lappaceum Hiern (1875) dùng thuật ngữ coccous (lá noãn nứt vách, quả hạch nhỏ); Lecomte (1912) dùng partie (phần, thùy); Leenhouts (1994) dùng lobe (thùy); Welzen (cùng Verheij, 1992) dùng schizocarp (quả nứt/ nẻ, liệt) nhưng vào năm 1999 ông lại dùng thuật ngữ lobe,... Chúng tôi thấy rằng, các phần bị xẻ ra đó được phát triển từ một phần của bầu (khác với phân quả ở Annonaceae - được phát triển từ 1 lá noãn đầy đủ), do đó đã đề nghị gọi 1 phần bị xẻ ra đó là giả phân quả (pseudofruitlet). Tuy nhiên, còn có một số ý kiến khác nhau về vấn đề này, vì thế trong công trình vẫn dùng thuật ngữ thùy (lobe) như đa số ý kiến đề nghị.
(1) Do cấu tạo rất đặc biệt của hoa đơn tính trong họ Bồ hòn (luôn có cả bộ nhị và bộ nhụy trên cùng một hoa, nhưng chỉ một trong hai bộ phận có chức năng sinh sản, bộ phận còn lại sẽ bị thui sau khi hoa nở), cho nên việc sử dụng thuật ngữ đối với những hoa kiểu này vẫn chưa thống nhất. Quan điểm thứ nhất cho rằng những hoa này hầu hết là hoa lưỡng tính, vì vậy đã sử dụng thuật ngữ tạp tính (polygamous, polygame, polygamus). Theo quan điểm này có Bentham & Hooker (1862), Hiern (1875), Pierre (1894-1895), Lecomte (1912),... Quan điểm thứ hai sử dụng thuật ngữ đơn tính (unisexual, unisexuel, unisexualis) và được đa số các nhà thực vật học sử dụng, như Radlkofer (1932), Adema (1994), Leenhouts (1969, 1971, 1994),... Gần đây, nhà thực vật Hà Lan Welzen (1997) đã sử dụng thuật ngữ staminate flower để chỉ hoa có chức năng của hoa đực và thuật ngữ pistillate flower để chỉ hoa có chức năng của hoa cái. Ở Việt Nam, qua nghiên cứu chúng tôi thấy nếu gọi những hoa đơn tính nêu trên là hoa lưỡng tính thì không đúng (vì những hoa này chỉ có một tính - đực hoặc cái), còn nếu gọi là hoa đơn tính thì cũng không thật chính xác (vì trên cùng một hoa luôn có cả bộ nhị và bộ nhụy), chính vì thế chúng tôi đã đề nghị gọi những hoa đó là hoa giả tạp tính (pseudomixis, spuriously polygamous). Tuy nhiên, sau khi dùng thuật ngữ này, chúng tôi thấy có nhiều ý kiến không ủng hộ. Gần đây, GS.TSKH. Trần Đình Lý đã đề nghị gọi những hoa đó là hoa lưỡng tính giả. Chúng tôi thấy rằng, để tránh những ý kiến trái ngược nhau như trong thời gian vừa qua cần có một hội thảo bàn về vấn đề này để có sự thống nhất rộng rãi. Chính vì vậy, trong công trình này, chúng tôi vẫn sử dụng thuật ngữ đơn tính như đa số ý kiến đề nghị.






_254903291020251.jpg)






